人类大脑活动的时空复杂性结构
原创

人类的大脑运作在大范围的功能网络中。这些网络是不同脑区域之间时间相关活动的表现,但全局网络特性和单个脑区神经动力学的关系仍然不完全清楚。本文展示了大脑的网络体系结构与神经正则性的关键时刻紧密相连,这些时刻表现为功能性磁共振成像信号中的自发“复杂性下降”,反应了脑区之间的功能连接强度,促进了神经活动模式的传播,并反映了个体之间的年龄和行为差异。此外,复杂性下降定义了神经活动状态,动态塑造了脑网络的连接强度、拓扑配置和层次结构,并全面解释了脑内已知的结构-功能关系。这些发现描绘了一种原则性的神经活动复杂性体系结构——人类的“复杂组”,它支撑着大脑的功能网络组织。
简介
人类的大脑运作在大规模功能网络中,这些网络总称为功能性人类连接组,空间分布不同的脑区域之间具有时间相关性的活动。神经信号的这种协方差通常称为功能连接(FC),从功能性磁共振成像(fMRI)中估计FC揭示了大脑的功能体系结构中的重要特征。首先,FC在大脑中不是均匀分布的,而是被组织成为休息状态下的网络(RSNs)功能子系统。其次,FC不是静态的,而是表现为动态波动,形成明显的时间网络状态。第三,在网络结构中,没有随机性,而是遵循一种有效拓扑结构来反映网络内的通讯构型。最后,大脑网络在功能上不是统一的,而是沿着从低阶单模到高阶跨模处理系统的一种主层次组织。
目前,理解人类连接组的结构、时间动态和层次仍是一个根本性挑战,需要一个解释性框架将全局网络结构与个体脑区域的局部活动相联系。FC 以及基于FC计算的网络属性都是根据区域信号之间的协方差关系定义的,但这种网络建构具有方向性(区域活动定义区域之间协方差,但不可反向计算),网络的全局特性最终必须根植于单个脑区域的神经动力学。因此,神经活动波动的可变性导致了定义网络的协方差结构,但全局网络结构如何与局部神经活动的可变性联系仍然不完全清楚。
基于此,本研究利用信息理论复杂性分析(i)将单个脑区的局部活动与网络的整体特性相关联,(Ⅱ)在标准化空间中表达神经波动的固有变异性,并且(iii)允许对神经变异性进行具有时间分辨率的记录,以捕捉个体信号内的时间动力学。该方法基于血氧水平依赖(BOLD)活动的符号编码,利用信号中的幅度信息和抽象模式的多样性来量化在特定时间点的不规则程度(即信号复杂度)。由此,我们获取了每个脑区的复杂度时间序列,通过神经变异的元表征捕获局部动态,并允许节点(即个别区域)和网络中的边(即区域间的功能连接)之间保持联系。
材料和方法
本研究利用HCP项目中的343名被试fmri静息态数据。每个被试连续两天每天2次,共收集4次静息态数据,其中两次作为主分析数据,两次作为保留数据来验证。未完整数据被剔除。在提取时间序列之前,对每个体素进行带通滤波(0.01~0.1Hz)和z评分。使用脑网络组(BNA)图谱提取区域BOLD时间序列,该图谱包括246个皮质和皮层下感兴趣区域(roi)。分配7个规范静息态脑网络(视觉,躯体运动、背侧注意、腹侧注意、默认模式、额顶叶和边缘叶)。基底神经节、丘脑、海马体和杏仁核中的区域被划分为皮层下区。本研究还计算了功能分割和时间序列提取的方法对复杂性动态选择的敏感性。最后,将BNA分割网络与两个高空间粒度和低空间粒度的体积图谱,以及360个皮质roi的表面基多模态分割(MMP)图谱进行了比较。
时间序列估计采用的是通过对时间序列向量进行符号编码,计算出BOLD活动的信号复杂度(WPE),WPE是排列熵(PE)对振幅敏感,是一个捕获模式程度的信息理论量。传播分析是图论分析方法,每个单独的传播被建模为一个有向图中的节点表示复杂性下降的大脑区域,有向边表示从一个BOLD窗口到下一个窗口的时间进程,采用了逐步搜索。首先,在时间分辨亲和向量中识别出所有至少n = 10区域同时显示复杂度下降的BOLD窗口。如果有几个时间上连续的窗口满足这个标准,则将下降区域数量最多的窗口定义为有向图中的峰值层。从这个峰值层中,窗口旁窗口向前后搜索识别相邻窗口,其中靠近峰值层下降区域的数量增加(传播阶段)而在峰值后下降(褪色阶段)。因此,最小的级联长度包括三个连续的窗口(初始化峰值衰减)。为了研究在传播阶段的复杂度下降,我们从初始化窗口构建了单个有向图,其中显示下降的区域代表图的源节点—到峰值窗口(图2B)。本算法利用R的图形图包(版本1.2.5)实现。
使用默认的完全连锁方法对平均距离矩阵进行层次聚类。通过定义单峰和跨峰聚类群,明确地研究了这种聚类结构,随后使用符号包使用蒙特卡罗模拟进行测试。复杂状态的聚类为无监督结构的K-means聚类,以调查时间差异的复杂性状态。我们指定了k=4(即两个中间复杂度状态),并对k=3(即一个中间状态)和k=5(即三个中间状态)进行了全面验证分析。为了评估个体参与者的经验状态访问是在k个可能的复杂性状态中分布的程度,定义了一个状态分散指数(SDI),SDI位于[0,1]中,如果参与者只访问过一个状态(最小状态探索),则边界为0,如果经验状态占用与统一状态相同,则边界为1(最大状态探索)。为了表征在估计的复杂性状态后的空间拓扑结构,定义了一个跨复杂性状态的距离指数(DACS),由此构成了后续相关分析的输入,将状态拓扑结构与皮质髓鞘形成和初级FC梯度以及结构功能耦合联系起来。
血氧波动是从边缘时间序列中估计脑区域间的BOLD共波动,它需要对两个z-得分的时间序列之间的相关性进行精确的数学分解,通过省略它们的元素级乘积的平均值的计算来实现的,从而产生了两个区域共波动的时间分辨向量,从框架水平解释相关性。同时计算平方和平方根,并将RSS时间序列阈值化,以获得高振幅波动事件。
功能连接和网络图谱分析包括静态功能连接(为整个静息状态记录上的roi-roi皮尔逊相关矩阵)和结构连通性矩阵(采用概率束状图进行计算)。引入的结构解耦指数估计结构-功能耦合(结构-函数解耦指数-SFDI)。SFDI量化了功能信号对解剖结构的依赖性。结构连接体是分解后的谐波分量,每个时间点的大脑活动都被写成谐波分量的加权线性组合,关键的步骤需要一个分裂的过程,将大脑活动分解为耦合和非耦合的部分,然后解耦合耦合信号规范的比例产生SFDI。
使用从t1加权/t2加权图像比值中获得的皮质灰度髓磷脂图来计算图谱模板中roi的平均髓磷脂含量。利用群平均FC矩阵计算宏观连通性梯度。然后应用了各自的皮质图的自旋排列相关性检验这些梯度负荷与其他皮质特征(髓鞘形成和DACS)之间的联系。观察到的这些皮质特征的关联进一步使用ppcorR包进行非参数偏相关分析。
为了评估信号复杂性和参与者特征之间的关联,将每个ROI和参与者的平均WPE值与年龄和认知表现、运动技能指数以及感官得分进行了分析,通过PLS分析BOLD信号复杂度与个体参与者测量之间的多变量关系。PLS通过计算一个相关矩阵来工作,该相关矩阵捕获了每个区域的目标大脑测量和感兴趣的行为指标的参与者之间的相关性。
结果
大脑活动具有神经规则性的关键时刻
人类连接体项目的静息态功能磁共振成像,揭示了人类大脑活动的一个高度典型的复杂性结构:超过的80%的采集时间内大脑表现出广泛的高复杂度信号,在所考虑的潜在的信号特征条件下,接近期望值上限。期间也有出现由于BOLD信号规则性上升导致的瞬时性“复杂度下降”,如图1A所示。在这里,典型的丘脑区域(第一行)的相对振幅变化在很大程度上被分为一个“向上”的基序(1-2-3)和一个“向下”的基序(3-2- 1)。相比之下,在左侧楔前叶,几乎所有在给定窗口上的振幅变化都归因于向下的基序,导致了一个高度规则的模式分布,因此,在这个窗口上的复杂性下降了。空间分析显示,内侧颞叶、前扣带和皮层下区域持续高复杂的活动,最显著的是丘脑区域(图1B)。相比之下,复杂性下降发生在大脑皮层,主要集中在中线周边区域。在个体水平上,复杂度下降接连性与大平均脑信号复杂度密切相关,在不同RSN 和年龄阶段间也存在显著差异。将单个区域的复杂性时间序列与这些区域之间的FC联系起来。任何两个区域之间的FC强度与它们表现出复杂性下降的程度密切相关(图1C)。虽然这种“下降巧合”解释了FC整体的大部分方差,但标准的RSN内部连接也显著增高(W=3.6×107,P = 9.3×10−161),高于跨脑区的网络连通性。区域复杂性时间序列的半球间对称性(图1D)进一步量化为每个区域与它的对侧等位置的秩加权相关性证明了网络分化。这种半球间的对称性受到典型的RSN[χ2(7)=169,P=4.6×10−33]的系统限制,并在本质上反映了最近发现的单模态和传输之间的层次结构,表明神经活动模式的跨半球耦合在初级网络中最强,并在高阶系统中逐渐变得更加多样化。
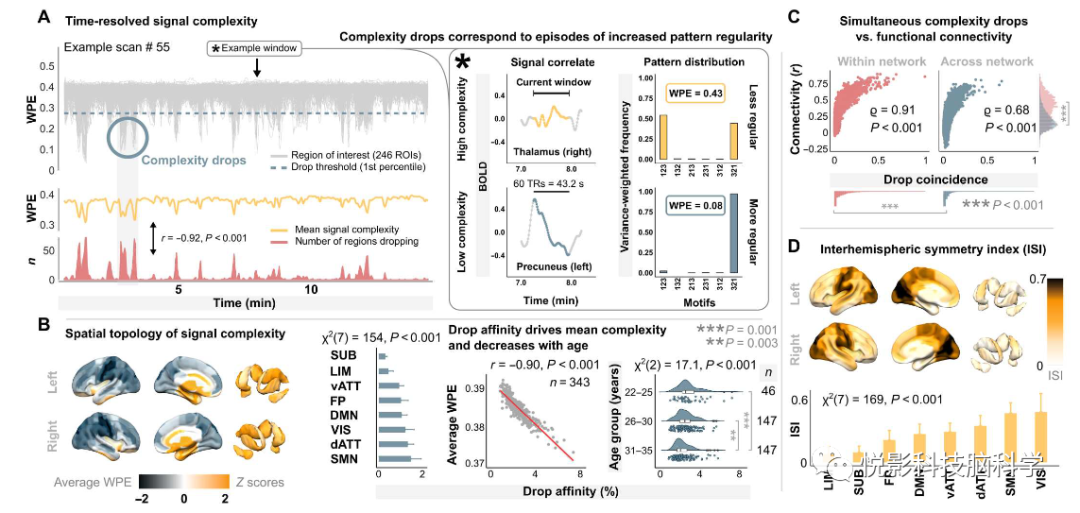
图1.在短暂的复杂性下降时可见规则性人类大脑活动的关键时刻。(A)对中每个ROI的神经信号的时间分辨WPE典型的静息状态记录(脑网分割;窗口长度为60 TRs,95%重叠)。插图显示了两个具有代表性的高复杂度和低复杂度信号的BOLD关联。(B)平均信号复杂度的空间分布。RSNs中下降区接近性与大平均信号复杂度和年龄相关。(C) 在典型RSN内和外FC作为下降次数的函数。(D)大脑半球间复杂性对称性的空间拓扑结构和网络分布。dATT、背侧注意、DMN、默认模式、FP、额顶叶、LIM、边缘、SMN,躯体运动;子下,皮层下;vATT、腹侧注意;VIS、视觉网络。
规则性时刻贯穿整个大脑
复杂性的下降始终发生在动态级联中,从一些初始化区域开始,逐渐扩散到整个大脑,最后再次消失到几个区域,随着时间的推移,全脑范围内的下降形式呈现出倒u形。如图2A所示,这种级联在所有343名参与者中都存在,通常持续约10秒。图2B利用图理论框架描述每个单独的传播的有向图。这些传播图包含节点层,这些节点层表示从级联(“源层”)初始化到最大下降区(“峰值层”),以及其中边的变化表示时间的进展,允许估计传播中区域到区域的转移概率。
下降级联反应的特征表现为:级联过程的持续时间与参与其中的区域数量之间有着强烈的正相关关系。单个体呈现的级联数量与他们的源多样性密切相关,定义为曾经初始化级联的独特大脑区域的比例(ρ = 0.96,Padj = 2×10−182;图2C)。值得注意的是,我们观察到与年龄相关的级联反应数量和源多样性显著减少,但没有在数量下降或级联持续时间中有显著变化,表明个体传播以半变异的方式发生(图2C)。对传播图的空间分析显示,下降级联主要起源于皮层,重点是低阶RSN(图2D)。此外,从个体扩散的转移概率构建的图揭示了一个高度结构化的时空传播网络(图2E)。网络的中心性分析表明复杂度下降在区域间的传播遵循一个内在的层次结构(图2F),其中影响最大的节点对应于主RSN,高阶系统的节点对传播的重要性逐渐降低。零模型检验显示复杂性下降沿着一种单峰到多峰功能梯度的时空路径传播(图2G)。这种传播结构再次强烈地与连接组内部的功能层级汇聚,特点是在低阶单峰和高阶多峰处理系统之间有主要梯度。将测地线距离建模为概率流线连通性和结构连接数的函数显著提高了解释力度,与只考虑链路数时相比,下降跨区传播进一步受到结构连通性的限制。
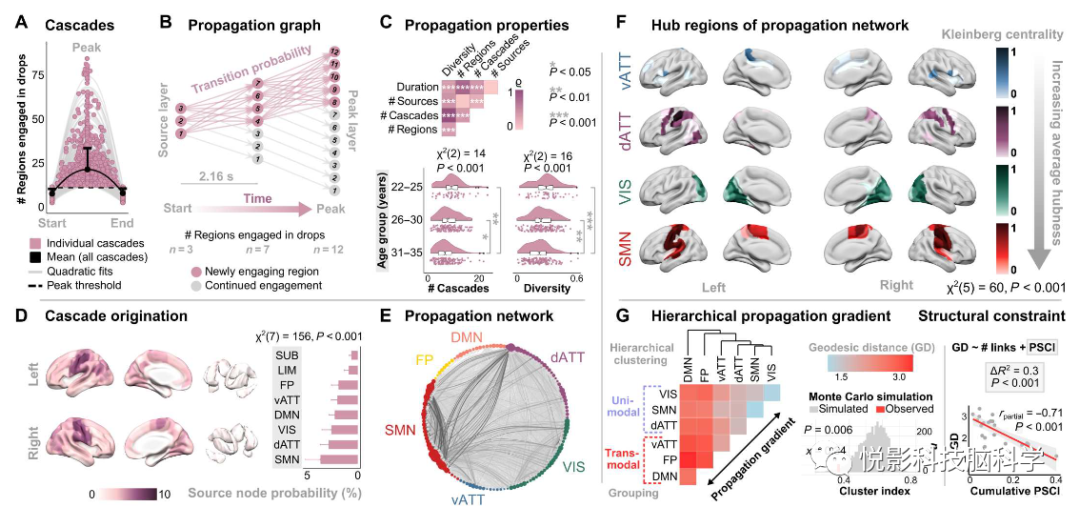
图2.复杂性下降沿着主要功能层次贯穿整个大脑。(A)下降级联。点代表了研究人群中5279个级联中的500个随机样本,级联的阈值为至少10个区域在峰值时同时下降。(B)个体有向传播形式图。窗口到窗口的时间分辨率关联应答为2.16秒(3 TRs)。(C)传播特性和年龄效应(成对fdr校正)。(D)级联起源的空间拓扑结构。(F)阈值传播网络的拓扑结构。FP 和DMN 因几乎零数据未显示。(G)传播网络中节点间的平均测地线距离。PSCI,概率流线连通性指数。
网络调控的复杂性状态组织神经活动
将单个区域的复杂性动态与全脑网络的行为联系起来,在复杂度时间序列上使用k-means聚类的无监督结构检测时序性脑网络状态,表明神经信号处于不同的时间网络状态(图3a),与个体时间序列的检查一致(图1a),参与者大部分时间处于高复杂度的默认神经状态,而访出现次数更多的状态需要逐渐降低复杂性,导致在不同时间下存在显著的复杂性区分效应(η2 = 0.90,[0.89-0.91])。值得注意的是,虽然复杂性状态直接来自单个大脑区域的活动,但它们对连接强度和拓扑产生了混淆作用。特别是,神经活动的默认状态特征是低强度的网络连接,而较低复杂度的状态导致连通性逐渐增加(图3a)。在个体水平,复杂性状态的网络调制效应依旧存在于时间窗内;群体水平,年龄较大的参与者表现出越来越严格的神经动力学,这与年龄相关的下降亲和力(图1B)和传播多样性(图2C)的降低相一致。值得注意的是,所有的状态相关结果对聚类参数定义的预期复杂性状态的数量具有高度鲁棒性。
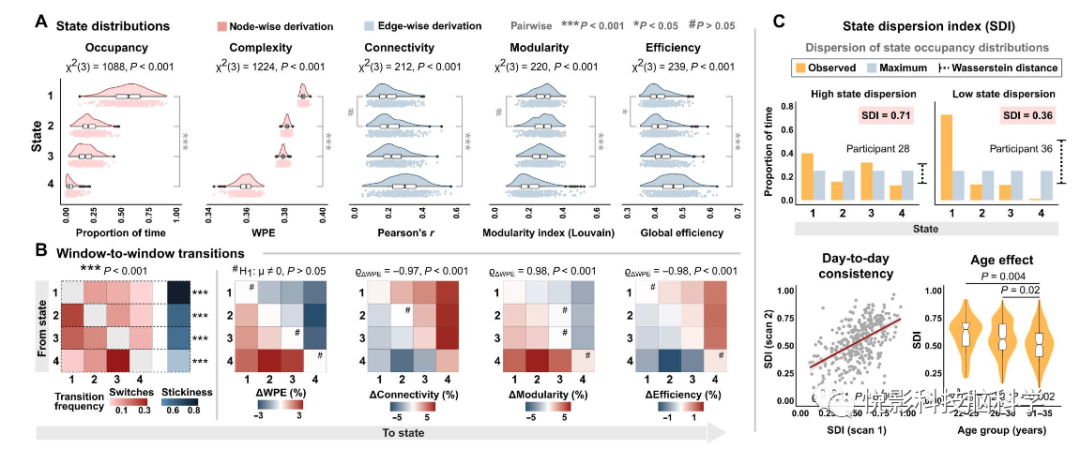
图3.神经活动具有调节网络强度和配置的时间复杂性状态。(A)占用率、信号复杂度、连通性强度的状态级分布,网络模块化,和全局效率。斑点代表参与者(n = 343)。节点推导参考单个区域信号的估计,边缘推导参照来自网络的关系(即区域到区域的相关性)。(B)窗口到窗口的状态转换频率。状态依赖的变化测试偏离零。网络度量与ΔWPE的关联对应于各自矩阵的相关性。(C)连续两天的一致性,并与年龄相关的降低状态分散指数(SDI)。
复杂性状态连接了结构和功能的网络层次
从FC数据中明确相应的梯度负荷,并将其与复杂性状态的拓扑联系起来,结果发现复杂度状态的拓扑结构证实了平均信号复杂度的分布(图1B),皮层下区域始终显示出高复杂度的活动,在很大程度上独立于当前全局复杂度状态(图4A)。相比之下,皮层拓扑结构随着复杂性下降的区域数量而明显变化,导致显著的空间异质性。定义一个跨复杂性状态的距离度量(DACS)为每个区域的累积质心到质心的欧氏距离跨越四维状态空间(图4B),分析结果表示,DACS拓扑结构密切遵循一个单峰到跨峰的梯度,其中在复杂状态中变化最大的区域代表了层次结构的单峰端。层次回归方法表明,复杂性拓扑结构单独解释了梯度负荷的大部分方差,并解释了髓鞘梯度效应的复合增强模型(图4D)。
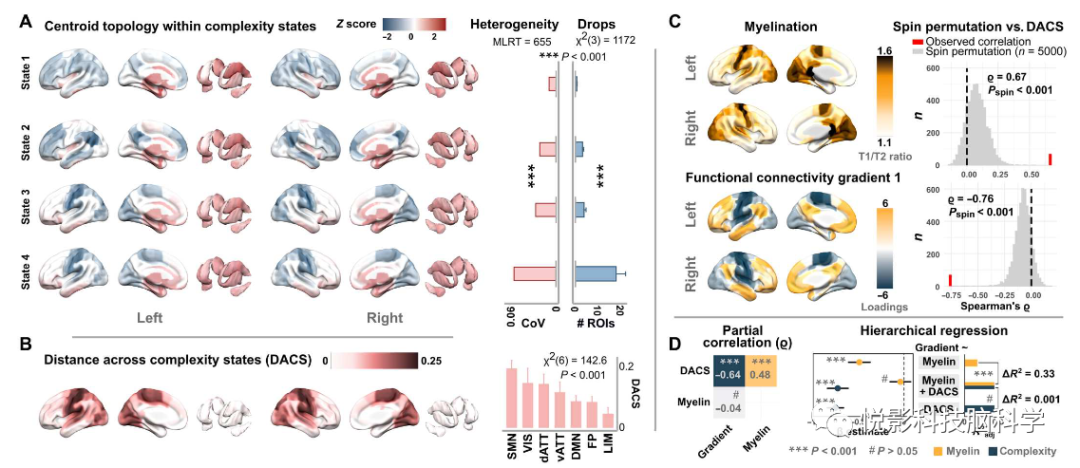
图4.复杂性状态的空间嵌入全面地解释了大脑内的结构-功能关系。(A)质心位置的空间拓扑结构(状态内z分数)、区域间的变异系数(CoV)和每个状态的平均复杂性下降数。(B)跨复杂度状态的距离的空间拓扑结构和网络分布(DACS)。(C)平均区域髓鞘化(以t1加权/t2加权图像比值代表)和主要的单峰到跨峰FC梯度与通过自旋排列试验评估的DACS的相关性。(D) 复杂性、髓磷脂和连通性梯度之间的偏相关性,以髓磷脂含量和DACS为解释变量的梯度负荷的层次回归。
复杂性状态反映了大脑网络中的结构-功能耦合
如图5A所示感觉-运动区域的结构-功能耦合更强,高认知区域的结构-功能耦合较低,这种分布与复杂性状态的空间拓扑结构联系起来,揭示了一个宏观皮层组织的多模态梯度(图5B),它连接了结构连通性、FC和局部结构神经动力学。在这个梯度中,一端包括显示许多复杂性下降的区域(在复杂性状态上高度可变),代表功能层次结构的单峰端,并显示高结构函数耦合。相比之下,相反的一端包括复杂性下降较少的区域(在复杂性状态上变化较小的区域)代表了功能层次的跨峰极点相位,并显示出较低的结构-功能耦合。
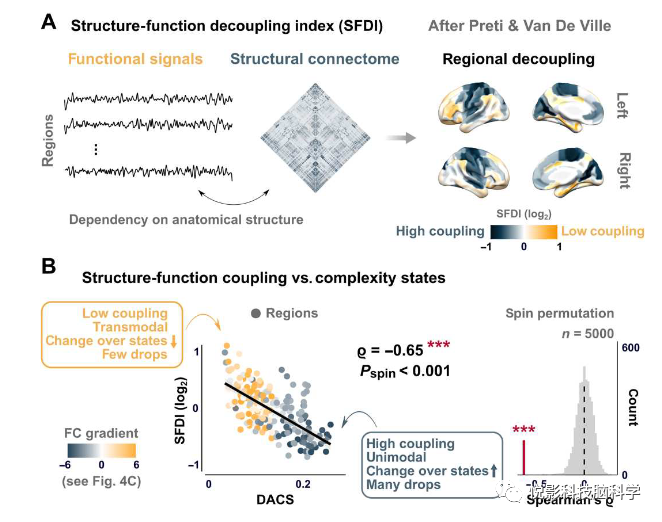
图5.复杂性状态反映的大脑网络中的结构-功能耦合。
复杂性动态在保留数据中具有高度一致性
对来自同一参与者的保留数据的验证分析显示,与主要分析相比信号复杂度的分布非常一致,产生几乎相同的下降阈值(图6A)。尽管是高度动态的指标,但个体参与者对复杂性和他们的复杂度的平均信号复杂度的亲和力在主数据和保留数据中具有很强的相关性(图6B)。对保留数据的独立聚类紧密地证实了神经复杂状态的分布,产生了一个主导的高复杂状态和逐渐更少见的低复杂状态(图6C)。此外,参与者探索复杂性状态的程度(由SDI测量)以及复杂性状态在大脑中的空间嵌入(由DACS测量)在保留数据中也高度一致(图6D)。
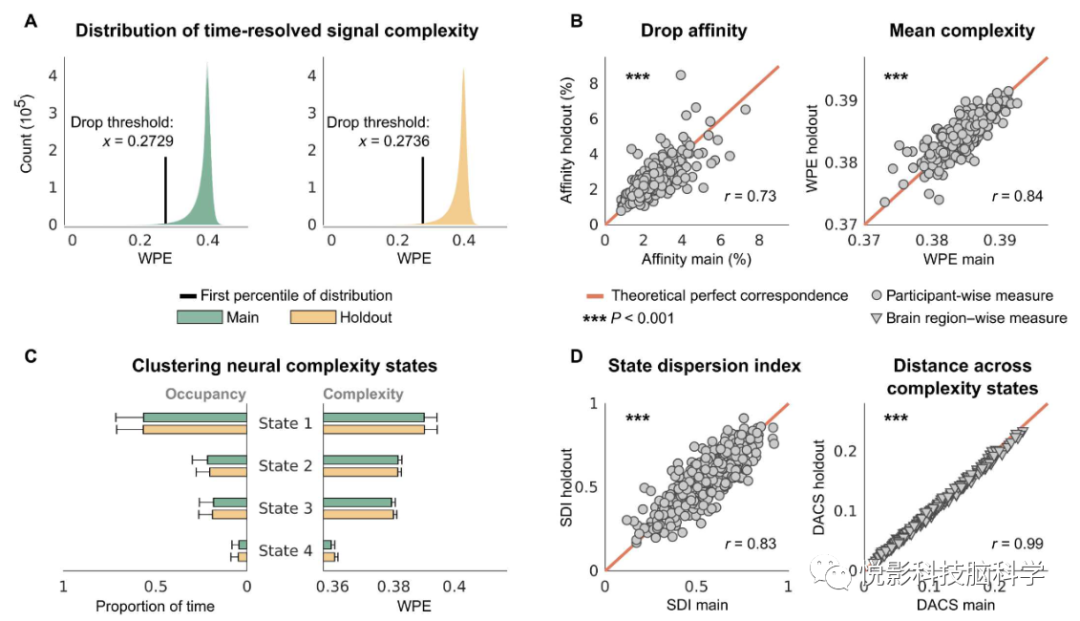
图6.在保留数据中复杂性测量高度一致性
复杂性行为关联
对主数据集和保留数据(图7)以及单个扫描session进行了这些分析。来估计行为的一致性。PLS分析返回了一个显著的复杂性差异和行为差异之间关系的潜在解。在定向期望中,潜在大脑得分与潜在行为得分呈正相关。在这个潜在空间中,神经复杂度为与年龄正相关,与流动和结晶能力、认知任务表现和运动功能呈负相关,在保留数据中具有高度一致性(图7、A和C)。因此,相关的引导比率(BSRs)表明,复杂性-行为关联受到功能网络的系统约束,与腹侧注意和默认模式网络相关的区域贡献最强,这种效应在主数据和保留数据中都可以观察到(图7、B和D)。
讨论
总的来说,这些发现描述了基于神经活动的时空复杂性结构的大脑组织统一原则,解释了神经信号的可变性和大脑功能网络的几个关键特性,对我们理解大规模的大脑动力学有五个直接启发:
1)静息状态的大脑活动的特征是神经规律性的关键时刻,复杂性的下降是自发的,在时间和空间上都是高度协调的,并紧密地解释了功能连接强度作为大脑区域同步程度的指示。但是,同时复杂度下降的区域常位于同一功能网络,在传统上被分配到不同功能网络的区域之间也有大量的下降巧合,这支持了大脑区域可以动态参与不同的时间内、不同的功能。
2)复杂性的下降将人类大脑活动的两个关键现象联系起来:神经模式的动态传播和大规模大脑网络的功能层次。观察到复杂性的下降通常可以从大脑的任何地方开始,它们沿着高度结构化的时空传播路径在大脑皮层扩散。
3)复杂性的下降定义了神经活动的时间状态,神经复杂性状态动态地调节了母网络的连接强度和拓扑配置一个时刻的形式。研究结果表明,大脑可能会实现与一个默认的神经状态的分离活动的平衡,以维持成本效益,而更罕见的低复杂度以确保功能集成的周期性阶段。
4)复杂性状态的空间嵌入全面解释了大脑中建立的结构-功能关系。
5)观察到的复杂性动态携带了许多个体间的功能含义,这些含义在主数据集、保留数据集和个体扫描会话中都是稳健的。考虑到个体的亲和力下降与整体信号的复杂性和年龄密切相关,复杂性的下降与功能整合的阶段紧密相关,以及腹侧信号的下降阶段密切相关的注意网络被认为是网络之间切换的中介系统,可见,更高的复杂度下降容量可能属于脑活动的有益表现。
本文仍存在一些技术性问题,首先是fMRI的数据的TR较短,导致空间分辨率低;其次是信号复杂性的空间分布显示,在功能磁共振成像记录中内侧颞叶和皮层下区域具有一致的高复杂度活动,其时间序列通常显示出较低的信噪比。虽然信噪比对复杂度估计的影响不能完全消除,但这些区域的高信号复杂度可能不是完全由噪声导致。最后是,在不同的分析下定义哪个空模型的问题依旧需要进一步探讨,看看经验复杂性动态是否与其他包含不同属性的替代方法有相关结果。
参考文献:A spatiotemporal complexity architecture of human brain activity.
原创声明:本文系作者授权腾讯云开发者社区发表,未经许可,不得转载。
如有侵权,请联系 cloudcommunity@tencent.com 删除。
原创声明:本文系作者授权腾讯云开发者社区发表,未经许可,不得转载。
如有侵权,请联系 cloudcommunity@tencent.com 删除。
评论
登录后参与评论
推荐阅读
腾讯云开发者

Copyright © 2013 - 2026 Tencent Cloud. All Rights Reserved. 腾讯云 版权所有
深圳市腾讯计算机系统有限公司 ICP备案/许可证号:粤B2-20090059 ![]() 粤公网安备44030502008569号
粤公网安备44030502008569号
腾讯云计算(北京)有限责任公司 京ICP证150476号 | 京ICP备11018762号